Желто зеленые водоросли примеры. Характеристика основных типов водорослей. Разнообразие и систематика
К отделу Желтозеленые водоросли относятся организмы, стоящие на разных ступенях морфологической дифференцировки таллома, - одноклеточные, колониальные и многоклеточные. Среди них встречаются преимущественно коккоидные, пальмеллоидные или нитчатые структуры, реже - амебоидные, монадные, сифональные, разнонитчатые и пластинчатые.
Для подвижных форм желто-зеленых водорослей (в том числе и для зооспор) характерны наличие двух неравных по размерам жгутиков (боковой - короткий, бичевидный и передний - длинный с мастигонемами) и желто-зеленая окраска хроматофоров, обусловленная наличием хлорофиллов а и с, каротинов (3 и £, ксантофиллов (антераксантин, лютеин, зеаксантин, вошериаксантин, виолаксантин и неоксантин). В зависимости от преобладания тех или иных пигментов встречаются виды со светло- или темно-желтой окраской, реже зеленой и у некоторых - голубой. Запасные продукты - волютин, жир, часто хризоламинарин. У примитивных форм содержимое клетки окружено тонким перипластом, а у более высокоорганизованных представителей имеется пектиновая или целлюлозная оболочка (цельная или двухстворчатая). Оболочка клеток часто пропитана солями железа, кремнеземом, известью, имеет различные «скульптурные украшения».
В протопласте клетки есть несколько хроматофоров, которые могут быть дисковидной, пластинчатой, ленто- или чашевидной или звездчатой формы. Ядро одно или много. У некоторых видов имеются пиреноиды. У подвижных форм отмечена стигма.
Желто-зеленые водоросли могут размножаться продольным делением клетки, распадением колоний или нитей на отдельные участки, а также зоо- или апланоспорами. Половой процесс (изо- или оогамия) известен у немногих. Для перенесения неблагоприятных условий у отдельных видов образуются цисты со слабоокремневшей двухстворчатой оболочкой.
Водоросли этого отдела встречаются главным образом в чистых пресноводных водоемах, реже в морях и солоноватых водах и почве.
Отдел включает два класса: Ксантофициевые (Xanthophyceae) и Эвстигматофициевые (Eustigmatophyceae).
Класс Ксантофициевые (Xanthophyceae )
Этот класс включает одноклеточные и многоклеточные организмы, преимущественно коккоидной структуры, реже наблюдается монадная, ризоподиальная, пальмеллоидная, нитчатая, разнонитчатая или сифоновая структура тела. Монадные формы и стадии с двумя неравными жгутиками и стигмой, расположенной на переднем крае хроматофора, под его оболочкой. Хроматофоры окружены каналом эндоплазматического ретикулюма, продолжающимся в наружную мембрану ядерной оболочки.
В соответствии с типами организации таллома класс делят на 6 порядков: Гетерохлоридальные (Heterochloridales ), Ризохло-ридальные (Rhizochloridales ), Гетероглеальные (Heterogloeales), Мисхококкальные (Mischococcales ), Ботридиадьные (Botrydiales) и Трибонематальные (Tribonematales).
Ниже рассматриваются 2 последних порядка.
Порядок Ботридиальные (Botridiales ). Для представителей порядка характерна сифональная структура таллома. Внешне они могут иметь сложную форму, но по строению протопласта представляют собой одну гигантскую многоядерную клетку. Как правило, таллом дифференцирован на окрашенную наземную и бесцветную подземную части.
Типичными представителями порядка являются роды ботридиум и вошерия.
Род ботридиум (Botrydium) объединяет наземные прикрепленные сифональные водоросли, имеющие шаро-, груше- или булавовидную форму. Подземная часть у них представляет дихотомически разветвленную систему бесцветных ризоидов. Клетка покрыта пектиновой оболочкой, которая, пропитываясь известью, с возрастом грубеет. В постенном слое цитоплазмы располагаются пластинчатые или дисковидные хроматофоры и многочисленные капли масла. Мелкие ядра видны только после окраски.
Размножается ботридиум зооспорами, иногда авто- или апланоспорами. При неблагоприятных условиях (продолжительное высыхание) содержимое наземной части (шарика) перемещается в ризоиды и распадается на отдельные части, покрытые толстой оболочкой, образуя покоящиеся цисты - ризоцисты. С наступлением благоприятных условий ризоцисты прорастают в новые особи непосредственно или через стадию зооспор.
Известно более 10 видов, в том числе 1 вид - Б. зернистый (В. granulatum ) - в Беларуси. Развиваются на глинисто-илистых отложениях берегов водоемов, на дне подсыхающих прудов, в колеях проселочных и лесных дорог, на влажных, богатых питательными веществами почвах с повышенным содержанием извести.
Род вошерия (Vaucheria) включает водоросли, сифональный таллом которых представляет собой неправильно и редко ветвящиеся нити нежного светло-зеленого цвета с бесцветными ветвящимися ризоидами. Это одна гигантская многоядерная клетка длиной несколько сантиметров. Центральную часть ее занимает крупная вакуоль с клеточным соком. В постенном слое цитоплазмы расположены многочисленные дисковидные хроматофоры без пиреноидов и капельки масла.
Бесполое размножение осуществляется многожгутиковыми и многоядерными зоо- и апланоспорами. При этом содержимое на концах ветвей становится более густым и темным, отчленяется перегородкой от общего слоевиша и превращается в зооспорангий, где формируется одна крупная зооспора, называемая синзооспорой, с многочисленными парными жгутиками по периферии.
Половой процесс у вошерии оогамный. На слоевище или на особых коротких ветвях образуются антеридий и около него один-два или несколько оогониев. При созревании яйцеклетки из носика оогония выступает капелька содержимого, привлекающего сперматозоиды. Один из них (с двумя жгутиками неравной длины) внедряется в оогоний через образоЕ!авшееся отверстие и оплодотворяет яйцеклетку. После оплодотворения в оогонии развивается зигота (ооспора) с толстой оболочкой, содержащая много масла и гематохрома. После периода покоя в ней происходит редукционное деление ядра и ооспора прорастает в новый таллом.
Известно 62 вида, распространенных по всему земному шару.
В других странах Европы встречаются В. сидячая (К sesillis), В. крючковидная (V. hamata), В. вильчатая (V. dichotoma) и др.
Многие виды вошерии широко распространены в различных пресных водоемах, на сильно увлажненной почве, немногие - в морях.
Порядок Трибонематальные (Tribonematales ). Объединяет формы, характеризующиеся нитчатой структурой таллома. Это наиболее высокоорганизованные представители желтозеленых водорослей. По внешнему виду они сходны с улотриксовыми из отдела Зеленые и многими видами из отдела Золотистые водоросли. Типичным представителем данного порядка является род трибонема.
Род трибонема (Tribonema) включает водоросли, нити которых неразветвлены. Вначале они прикреплены к какому-либо субстрату с помощью базальной клетки, затем вследствие ее отмирания всплывают на поверхность водоема и встречаются уже как свободноплавающие, образуя тину желто-зеленого цвета.
Характерным признаком, по которому нити трибонемы легко отличить от других нитчатых водорослей, является своеобразное очертание их концов в виде двух рожек. Это связано с тем, что оболочка клеток у трибонемы крепкая, двухстворчатая и состоит из двух одинаковых половинок. Край одной половинки находит на край другой посередине клетки. При делении клетки из средней ее части формируется цилиндрический участок новой оболочки, в котором закладывается поперечная перегородка. Таким образом половинки соседних клеток прочно соединены друг с другом и при разрыве нити на части или ее распаде на отдельные клетки образуются характерные Я-образные фрагменты оболочки. Хроматофоров в каждой клетке трибонемы обычно несколько, дисковидной формы, без пиреноидов.
Известно 22 вида трибонем, из них 6 в Беларуси. Распространены преимущественно в прибрежной зоне различных водоемов на водных растениях, камнях, отдельные - в почве; часто образуют мягкие ватообразные, неослизненные, желто-зеленые дернинки. Повсеместно встречаются Т. обыкновенная (Т. vulgare), Т. зеленая (Т. viride), Т. меньшая (Т. minus), Т. ровная (Т. aequale) и др.тая и состоит из двух одинаковых половинок. Край одной половинки находит на край другой посередине клетки. При делении клетки из средней ее части формируется цилиндрический участок новой оболочки, в котором закладывается поперечная перегородка. Таким образом половинки соседних клеток прочно соединены друг с другом и при разрыве нити на части или ее распаде на отдельные клетки образуются характерные Я-образные фрагменты оболочки. Хроматофоров в каждой клетке трибонемы обычно несколько, дисковидной формы, без пиреноидов.
При размножении в клетках образуются одна или две разножгутиковые зооспоры или апланоспоры, при выходе которых створки расходятся и нить водоросли распадается. Для перенесения неблагоприятных условий служат акинеты с толстой клеточной стенкой или цисты.
Известно 22 вида трибонем, из них 6 в Беларуси. Распространены преимущественно в прибрежной зоне различных водоемов на водных растениях, камнях, отдельные - в почве; часто образуют мягкие ватообразные, неослизненные, желто-зеленые дернинки. Повсеместно встречаются Т. обыкновенная (Т. vulgare), Т. зеленая (Т. viride), Т. меньшая (Т. minus), Т. ровная (Т. aequale) и др.
скольких миллиметров до нескольких метров (иногда до 60 м и более). Таллом нарастает в результате интеркалярного роста или за счет деятельности верхушечной клетки. По внешнему виду - это ветвистые кустики, корочки, пластинки, шнуры, ленты, сложно расчлененные на стебле, и листовидные органы. Слоевища некоторых крупных представителей имеют воздушные пузыри, удерживающие ветви в воде в вертикальном положении. Для прикрепления к грунту служат ризоиды или дисковидное разрастание в основании слоевища - базальный диск.
По морфологической и анатомической дифференцировке таллома бурые водоросли стоят на более высоком уровне, чем все другие группы. Среди них не известны ни одноклеточные, пи колониальные формы, ни талломы в виде простой неразветвленной нити. Таллом самых простейших из ныне живущих бурых водорослей гетеротрихальный, у громадного же большинства талломы ложно- или истиннотканевого строения (выделяют ассимиляционные, запасающие, механические, проводящие ткани).
Оболочка клеток снаружи ослизненная, состоит из пектиновых веществ и внутреннего целлюлозного слоя. Слизь защищает клетки от механических воздействий, пересыханий во время отлива и т.п.
В цитоплазме находится одно ядро и хроматофоры дисковидной, реже лентовидной или пластинчатой формы, вакуоли, у многих видов пиреноиды. Хроматофоры клеток бурых водорослей содержат хлорофиллы а и с, каротины и несколько ксантофиллов: фукоксантин, виолаксантин, антераксантин и (еаксантин. Эти пигменты определяют бурую окраску водорослей. Продуктами запаса являются полисахарид ламинарин, шестиатомный спирт маннит и липиды.
У бурых водорослей встречаются обе формы размножения: бесполое и половое.
Бесполое размножение осуществляется участками таллома. У некоторых водорослей имеются специализированные веточки (выводковые почки), которые легко отделяются и дают новые растения.
Кроме того, у большинства бурых водорослей бесполое размножение происходит посредством зооспор, у отдельных представителей - тетраспор и у единичных видов - моноспор. Зооспоры развиваются в одно- или многогнездных спорангиях. Образованию спор предшествует мейоз (исключениесоставляют циклоспорофициевые, у которых мейоз происходит перед формированием гамет).
Половой процесс изо-, гетеро- и оогамный. При изо- или гетерогамии гаметы образуются в многогнездных, многокамерных гаметангиях, которые могут развиваться из одной или многих клеток.
У наиболее высокоорганизованных бурых водорослей половой процесс оогамный. Яйцеклетка оплодотворяется вне оогония. Зигота без периода покоя прорастает в диплоидную водоросль.
Для большинства бурых водорослей характерна смена форм развития: для одних - изоморфная, для других - гетероморфная. Эти разные типы жизненного цикла ранее были положены в основу деления отдела Бурые водоросли на 3 класса: изогенератные с изоморфной сменой поколений, гетерогенератные с гетероморфной сменой поколений и циклоспоровые с одним порядком Фукусовые, где чередование поколений отсутствует. Однако деление бурых водорослей на изогенератные и гетерогенератные довольно условно, поскольку в обоих классах существуют представители с противоположным типом смены форм развития. Поэтому более правильным подходом к классификации бурых водорослей считается дедение их на 2 класса - Фэозооспорофициевые (Phaeozoosporophyceae ) и Циклоспорофициевые (Cyclosporophyceae).
Бурые водоросли почти все живут в морях как донные, эпифитные или планктонные организмы. Заросли бурых водорослей являются пищей, местом размножения и укрытия многих видов животных, субстратом для микро- и макроорганизмов, одним из основных источников органики в умеренных и приполярных широтах.
Они применяются в промышленности (пищевая, парфюмерная, текстильная) благодаря наличию таких ценных веществ, как альгиновая кислота, альгинат, маннит и др.
Класс Фэозооспорофициевые (Phaeozoosporophyceae)
Для большинства водорослей класса Фэозооспорофициевые характерны 2 самостоятельные формы развития - спорофит и гаметофит или гаметоспорофит, которые могут быть сходными по внешнему виду, строению и размерам и разными, т.е. имеется - изоморфная и гетероморфная смена форм развития. У примитивных представителей смена форм развития отсутствует.
Класс Фэозооспорофициевых делят на 11 порядков, из которых ниже приводятся 5.
Порядок Эктокарпальные (Ectocaгрales ). Включает бурые водоросли, слоевища (как спорофита, так и гаметофита) которых построены из однорядных нитей, способных ветвиться. Размеры их варьируют от микроскопических до 30 и более сантиметров. Эти водоросли образуют налет или кустики на скалах либо на других водорослях. Размножаются бесполым и половым путями. Органами размножения служат одно- и мно- гогнездные вместилища. Одногнездные всегда являются спорангиями, а многогнездные могут функционировать и как гаметангии.
Эктокарпальные встречаются во всех морях мира, предпочитая холодные воды Севера.
Виды рода эктокирпус (Ectocaгрus) имеют кустистое слоевище высотой 0,1 - 30 см. Оно состоит из тонких однорядных стелющихся и ветвящихся вертикальных нитей. Рост нитей интеркалярный или диффузный. Прикрепление к субстрату осуществляется ризоидами, которые у крупных экземпляров образуют своеобразную кору у основания ветвей. К вершине ветвей клетки сужаются и заканчиваются длинным бесцветным волоском.
Спорангии и гаметангии располагаются как боковые выросты ветвей. Внутри одногнездных спорангиев происходят
мейоз и митоз с последующим формированием двухжгутиковых зооспор. Зооспоры вырастают в гаплоидные раздельнополые организмы с многогнездными гаметангиями. Гаметы изоморфные, но отличаются поведением: женская теряет подвижность и выделяет пахучее вещество, привлекающее мужские гаметы, одна из которых оплодотворяет ее. Зигота без периода покоя прорастает в диплоидный спорофит.
В зависимости от экологических условий описанный цикл развития может усложняться. Так, на спорофите кроме обычных одноклеточных зооспорангиев могут образовываться и многогнездные - нейтральные, где формируются (без редукционного деления ядра) зооспоры - нейтральные споры. Такие диплоидные зооспоры дают начало новому поколению спорофитов. Иногда и гаметы ведут себя как зооспоры - при их партеногенетическом прорастании (без оплодотворения) развивается гаплоидная особь. Наконец, известно, что у одного из видов эктокарпуса (Е. confervoides ) с гетероморфной сменой форм развития гаплоидные зооспоры функционируют как гаметы, т.е. копулируют.
По внешнему виду, строению и распространению эктокарпус похож (различие лишь в деталях структуры органов размножения) на пилайеллу (Pylaiella). Эти наиболее простые бурые водоросли обычно живут в прибрежной зоне морей. Они часто встречаются в Белом, Баренцовом и Черном морях, где поселяются на камнях или талломах других крупных водорослей.
Порядок Сфацеляриалыше (Sp / iacelariales ). Включает водоросли с жесткими кустистыми слоевищами высотой от нескольких миллиметров до 30 см; ветви цилиндрические. В отличие от других бурых водорослей у сфацеляриальных каждая ветвь оканчивается крупной клеткой, за счет деления которой водоросли имеют строго верхушечный рост. Для их слоевища характерно основание в виде корковидной пластинки из нескольких слоев клеток.
Вегетативное размножение происходит посредством столонов (нитей из нескольких рядов клеток, стелющихся по грунту) или специальных выводковых почек, отделяющихся от ветвей.
У сфацеляриальных существует изоморфная смена форм развития.
Водоросли рода сфацелярия (Sphacelaria) встречаются во всех морях. Слоевище у его представителей имеет вид кустика высотой до 4 см, состоящего из пластинчатой подошвы и отходящих от нее разветвленных нитей. Каждое разветвление нити на вершине несет крупную клетку, которая делится только в поперечном направлении и обусловливает нарастание таллома в длину. Отчленяющиеся таким образом клетки в дальнейшем делятся в продольном направлении, благодаря чему образуются узкие клетки, и таллом становится многослойным и внешне состоящим как бы из члеников.
Порядок Кутлериальные (Cutleriales ). Включает бурые водоросли, для которых характерно трихоталлическое строение слоевища за счет зоны роста, располагающейся в базальной части многоклеточных волосков, которые находятся на краях пластинчатого слоевища или на вершине ветвей кустистого таллома. Клетки зоны роста делятся, отчленяя клетки в сторону периферии и к слоевищу. Периферические клетки обеспечивают рост волосков по мере их старения и отмирания, а слоевищные соединяются в плотные паренхиматические пластинчатые образования талломов. С возрастом слоевище дифференцируется на более мелкие окрашенные клетки коры и крупные бесцветные клетки сердцевины.
Этим водорослям свойственна изо- и гетероморфная смена форм развития. Половой процесс у них гетерогамный. При этом морфологически различны не только гаметы, но и гаметангии: женские имеют крупные камеры и меньшее количество гамет, мужские содержат множество гамет в мелких гнездах.
Род кутлерия (Cutleria) служит классическим примером гетероморфной смены форм развития. У гаметофита слоевище однолетнее, вертикальное, вееровидное или кустистое, высотой до 15 см. Гаметангии развиваются на разветвлен-
пых или простых однорядных выростах. Слоевище спорофита одно- или многолетнее, корковидное, диаметром до 10 см. В спорангиях образуется от 8 до 32 зооспор. Морфологически зооспоры одинаковы, но из них развиваются раздельнополые женские и мужские гаметофиты в виде кустиков. На женских гаметофитах в макрогаметангиях возникают макрогаметы, на мужских в микрогаметангиях формируются 64 микрогаметы. После оплодотворения зигота без периода покоя прорастает в спорофит.
Характер цикла развития кутлерии зависит от экологических условий. У южных берегов Великобритании гаметофиты лучше всего растут летом, а в Средиземном море около Неаполя - зимой и весной. У прибрежной Скандинавии и северной Великобритании развиваются лишь спорофиты, а в Средиземном море - гаметофиты. В Черном море единственный вид К., усыпанная (С. adspersa) представлен только спорофитами.
Порядок Диктиотальные (Dictyotales ). Объединяет виды, которые характеризуются апикальным ростом и обычно дихотомическим ветвлением в одной плоскости. Бесполое размножение посредством апланоспор (тетраспор). Половой процесс оогамный. Смена форм развития изоморфная.
Большинство диктиотальных растут в тропических и субтропических морях. Довольно часто они встречаются в Черном (виды родов диктиота, дилофус и падина) и Японском (диктиота) морях.
Виды рода диктиота (Dictyota) характеризуются вильчато-разветвленным слоевищем с плоскими, обычно расположенными в одной плоскости ветвями без продольного ребра. Таллом развивается из цилиндрического ризома, прикрепленного к субстрату ризоидами. Вершина каждой ветви оканчивается одной крупной клеткой. Внутри ветвей находится слой крупных бесцветных клеток, окруженный снаружи корой из одного слоя мелких интенсивно окрашенных клеток.
На спорофитах из поверхностных коровых клеток развиваются сорусы одногнездных спорангиев, где образуется по четыре неподвижные тетраспоры. Тетраспоры прорастают в гаметофиты. Диктиота - двудомная водоросль: на женских гаметофитах формируются сорусы одногнездных оогониев с одной яйцеклеткой в каждом. Антеридии образуются на мужских гаметофитах. Яйцеклетки выпадают из оогония и в воде оплодотворяются сперматозоидами. Зигота сразу же прорастает в новый организм - спорофит. Наиболее широко распространена диктиота дихотомическая (D. dichotoma).
Род дилофус (Dilophus) сходен с диктиотой, только у его представителей внутри ветвей имеется не один, а два и более слоев бесцветных клеток.
Род падина (Padina) включает виды с плоским, вееровидным слоевищем, целым или с несколькими вертикальными разрывами. На поверхности хорошо выражены концентрические полосы, соответствующие рядам волосков. Наружные слои падины представляют собой мелкоклеточную кору, срединная же часть таллома составлена из крупных бесцветных клеток. Цикл развития падины такой же, как и у диктиоты, с той лишь разницей, что падина однодомная: оогонии и антеридии образуются на одном и том же организме.
Наиболее известна П. павлинья (P . pavonia). Этот вид встречается у берегов Южной Европы и у атлантических берегов Центральной Америки, а также в Черном море.
Порядок Ламинариальные (Laminariales ). Ламинариальные являются самыми совершенными водорослями среди водорослей всех отделов. У них наиболее крупные спорофиты (от 0,5 до 60 м и более). По форме многие виды напоминают высшие растения. У большинства видов таллом имеет сложное анатомическое строение с дифференцировкой на ткани. Он расчленен на листо-, стебле- и корневидные части. Рост слоевища у ламинаричьных интеркалярный (за счет деления клеток меристемы у основания листовых пластин и на вершине ствола).
Ламинариальные бывают одно- и многолетними. У ряда многолетних видов листовидные пластины ежегодно сменяются новыми, а стеблевидные и ризоидальные образования функционируют несколько лет. Перед сменой листовых пластин происходит спорообразование. Из спор формируются мужские и женские гаметофиты. Половой процесс у них оогамный. Обитают ламинариальные главным образом в холодных водах Южного и особенно Северного полушарий, предпочитая места с сильным течением или прибоем, что обеспечивает им интенсивное поступление питательных веществ.
Род ламинария (Laminaria) включает виды, таллом которых расчленен на листовидную пластинку, ствол и ризоиды. Листовидные пластины ровные или морщинистые, цельные или рассеченные. Ствол и ризоиды многолетние, листовидная пластина меняется ежегодно. На продольных разрезах с черешка и органов прикрепления выявляется их достаточно сложное анатомическое строение. Наружная часть черешка являет собой кору, состоящую из нескольких слоев клеток с хроматофорами; промежуточный слой представлен крупноклеточной запасающей тканью и, наконец, внутренний (сердцевина) - проводящей и механической. Проводящая система включает в себя трубчатые нити с воронковидными расширениями в местах клеточных перегородок. Эти перегородки имеют поры и называются ситовидными пластинами , а нити - ситовидными трубками. В толщину черешок растет за счет деления клеток коры, которое происходит периодически, вследствие чего на поперечном разрезе черешка хорошо заметны концентрические слои, напоминающие годичные кольца высших растений.
При размножении на поверхности листовидной пластинки из коровых клеток группами (сорусами) образуются одногнездные зооспорангии, в каждом из которых формируется от 16 до 128 двухжгутиковых зооспор. В благоприятных условиях зооспоры прорастают в микроскопически мелкие нитчатые заростки - мужские и женские гаметофиты.
Половой процесс у ламинарии оогамный. Созревшая яйцеклетка выходит из оогония и закрепляется на его верхнем конце. В таком положении происходит оплодотворение. Зигота без периода покоя прорастает в спорофит. Женский гаметофит обеспечивает не только формирование половых клеток, но и место прикрепления будущему спорофиту.
Известно около 30 видов, распространенных преимущественно в морях Северного полушария. Ламинария японская (L. japonica), или морская капуста, - самый ценный промысловый вид среди бурых водорослей. Ее используют для приготовления салатов, консервов, первых, вторых и третьих блюд. В ряде стран, в том числе и в России (на Дальнем Востоке), ее культивируют.
Класс Циклоспорофициевые ( Cyclosporophyceae )
Класс Циклоспорофициевые объединяет водоросли, у которых в цикле развития отсутствует чередование поколений. Их диплоидные талломы несут только органы полового размножения, развивающиеся в специальных округлых вместилищах - концептакулах, или скафидиях. Мейоз у циклоспоровых происходит перед образованием гамет.
Бесполое размножение спорами отсутствует.
Все циклоспорофициевые - крупные водоросли. Слоевища у них дифференцированы на ткани: меристодерму, кору, промежуточный слой и сердцевину; ситовидные трубки отсутствуют.
Порядок Фукальные ( Fucales ). Объединяет водоросли, которые характеризуются кустистой формой слоевища с верхушечным ростом. Клетки осевых частей слоевища делятся слабо. Они вытянуты в длину и составляют сердцевину. На талломах ряда фукальных развиваются стерильные крипто- и цекостомы (ямки с волосками или полости), располагающиеся под поверхностью слоевища. Они служат главным образом для поглощения питательных веществ.
Род фукус (Fucus) включает виды с плоским ремневидным дихотомически разветвленным талломом длиной до 1 м. Вдоль лопастей таллома с гладкими или зазубренными краями проходит срединная жилка, переходящая в нижней части в черешок, который прикрепляется к субстрату базальным диском. У некоторых видов фукуса по бокам от средней жилки расположены воздушные пузыри в виде вздутий. Таллом нарастает за счет деятельности верхушечных клеток. При размножении концы таллома вздуваются, принимают светлую желто-оранжевую окраску и превращаются в рецептакулы, на которых образуются скафидии с отверстиями. Между парафизами на стенках женского скафидия формируются оогонии, мужского - антеридии. В оогонии фукуса образуется 8 яйцеклеток, в антеридиях - 64 сперматозоида. Оплодотворение происходит в воде, и зигота, в отличие от ламинарии, не связана с оогонием. Зигота прорастает без периода покоя.
Виды фукуса распространены у берегов холодных и умеренных морей Северного полушария, часто образуя большие заросли на литорали, что облегчает их сбор и использование. Виды фукуса применяют в качестве удобрений, корма для скота, производства кормовой муки, альгинатов и других химических веществ. В морях России встречается 5 видов этого рода. Наиболее известны Ф. пузырчатый (F. vesiculosus ) и Ф. двусторонний (F. distichus).
Род аскофиллум (Ascophyllum) включает один вид - А. узловатый (A . nodosum). Его слоевище, достигающее 1 - 1,5 м в длину, узкое, без средней жилки, с длинным, дихотомически ветвящимся главным побегом, на котором попеременно или супротивно расположены более короткие ветви, заканчивающиеся прозрачными ягодообразными вздутиями (рецептакулами), несущими скафидии. Слоевища раздельнополые. Мужские экземпляры - более темной окраски, с менее округлыми ветвями и более мелкими рецептакулами. Строение скафидиев аскофиллума сходно с таковыми у фукусов. В оогониях формируются четыре яйцеклетки.
Водоросли растут в нижнем, отчасти в среднем, горизонте литорали и в верхней сублиторали (в Белом море), а также распространены у берегов Азии, Европы и Америки.
У видов рода цистозейра (Cystoseira) слоевище кустистое, прикрепляется к грунту диском (редко ризоидами) и достигает в длину 2 - 6 м. Оно прямостоячее, в форме цилиндрических или многогранных осевых стволов с боковыми «листьями» или без них, у основания часто сплющенное. Воздушные пузыри однокамерные, без поперечных перегородок, располагаются в верхних частях таллома цепочками или беспорядочно. Скафидии обычно обоеполые: в полости, у их основания, развиваются оогонии с одной яйцеклеткой, на стенках, ближе к отверстию скафидия, формируются антеридии.
Многие виды рода цистозейра произрастают в Черном, Японском и Охотском морях и у Курильских островов. В Черном и Средиземном морях растут Ц. косматая (С. crinita) и Ц. бородатая (С. barbata), в дальневосточных морях на глубине до 25 м - Ц. толстоногая (С. crassipe). Ц. косматая выделяется среди других видов, произрастающих в морях России, тем, что от диска, которым слоевище прикрепляется к грунту, отходит до 20 вертикальных стволов высотой до 0,5 - 1,2 м, а у Ц. бородатой - один высокий ствол, который у старых экземпляров становится шероховатым.
Желто-зеленые водоросли широко
распространены по всему земному шару, особенно в
чистых пресных водоемах. Но встречаются они и в
солоноватых, кислых и щелочных водах, а также
обычны в почве. Большинство желто-зеленых
водорослей – планктонные организмы. Часто их
можно найти в скоплениях нитчатых водорослей и
среди зарослей высших водных растений у берегов
рек, прудов, озер.
Многие из желто-зеленых водорослей одно время
относили к отделу зеленых – из-за некоторого
сходства с ними. Однако желто-зеленые отличаются
от зеленых водорослей важными признаками: их
подвижные формы имеют жгутики разной длины и
строения. Главный жгутик состоит из оси и
перисторасположенных на ней мерцательных
волосков, а боковой жгутик короткий, гладкий.
Именно из-за этой особенности желто-зеленые
водоросли называют еще разножгутиковыми. У
подвижных форм у переднего конца хроматофора
имеется красный глазок.
В хлоропластах желто-зеленых водорослей
содержатся хлорофиллы a
и c
, a- и b-каротин и
три вида пигмента ксантофилла: лютеин,
виолаксантин и неоксантин. В зависимости от
комбинаций этих пигментов клетки окрашены в
светло- или темно-желтый цвет, реже – в зеленый, и
у некоторых – в голубой. Крахмал в клетках
желто-зеленых водорослей не образуется. Вместо
него накапливается масло, а у некоторых видов –
углеводы хризоламинарин и волютин. Оболочки
клеток этих водорослей разнообразны – от тонкой,
позволяющей клетке образовывать псевдоподии, до
плотной целлюлозной, цельной или двустворчатой,
часто инкрустированной известью, кремнеземом
или солями железа.
Вегетативное размножение у желто-зеленых
водорослей осуществляется продольным делением
клетки, бесполое – зооспорами и апланоспорами.
Половой процесс – изо- или оогамия – известен у
немногих видов.
1 – ботридиум; 2 – перидиниум; 3 – цератиум
Ботридиопсис эрийский
(Botrydiopsis
eriensis
) – представитель желто-зеленых
водорослей, одноклеточный организм, широко
распространенный по всему земному шару. Обитает
в почве, на дне прибрежной зоны водоемов, среди
зарослей, в биологической пленке биофильтров
очистных станций. Встречается обычно в небольших
количествах, но иногда, при массовом развитии,
придает воде или поверхности субстрата нежную
желто-зеленую окраску.
Ботридиопсис легко обнаружить, взяв во время
экскурсии в лес, на луг, на реку щепотку любой
почвы. Ее надо поместить в чашку Петри, увлажнить
и положить на ее поверхность стерильные
покровные стекла, на которых водоросли хорошо
разрастаются. Чашки выдерживают на свету при
комнатной температуре и по мере роста водорослей
на стеклах просматривают под микроскопом. Клетки
ботридиопсиса, которые можно увидеть на стекле,
могут быть молодыми, мелкими, диаметром около 6
мкм, с двумя хлоропластами, или взрослыми,
крупными, диаметром до 18 мкм, с двухконтурной,
иногда желтоватой оболочкой и со множеством
хлоропластов. Можно увидеть и клетки,
наполненные автоспорами и покрытые разбухшей
ослизненной оболочкой.
Миксохлорис (Myxochloris ) – желто-зеленая водоросль, которая обитает в клетках листьев сфагновых мхов. Ее тело представляет собой крупный многоядерный плазмодий. Осенью миксохлорис образует цисты, из которых весной выходят зооспоры или амебы, проникающие в пустые клетки листьев мха и сливающиеся там в плазмодий.
Трибонема зеленая
(Tribonema viride
) –
нитчатая желто-зеленая водоросль, широко
распространенная в различных водоемах. Нити ее,
собранные в ватообразные дерновинки, образуют в
прибрежной полосе желто-зеленые скопления, на
ощупь мягкие, но не ослизненные. Молодые нити
трибонемы долго остаются прикрепленными к
субстрату с помощью базальной клетки, но позже
она отмирает; нити отрываются и образуют
скопления, плавающие на поверхности воды или
опускающиеся на дно водоемов.
Под микроскопом трибонему легко отличить от
других нитчатых водорослей по характерному
очертанию концов ее нитей. Поскольку оболочка
клеток трибонемы крепкая, двустворчатая, разрыв
нитей при бесполом размножении происходит в
середине клеток. При этом зооспоры выходят
наружу, а оставшаяся оболочка приобретает вид
двух рожек на каждой краевой клетке. При
наступлении неблагоприятных условий в нитях
трибонемы образуются апланоспоры с толстой
двустворчатой оболочкой, заполненные каплями
масла и жира.
Ботридиум
(Botrydium
). Летом на
влажной почве берегов водоемов, по колеям лесных
дорог, по краям засыхающих луж можно увидеть
множество темно-зеленых блестящих пузырьков 1–2
мм в диаметре, напоминающих рассыпанный бисер.
Каждый такой пузырек – клетка желто-зеленой
водоросли ботридиума. Книзу клетка постепенно
сужается и переходит в ветвящиеся бесцветные
ризоиды, погруженные в почву. В центре клетки
имеется вакуоль, в пристенном слое цитоплазмы
находятся мелкие ядра, многочисленные
хроматофоры и капли масла. Считают, что такой тип
организации таллома является сифональным, или
неклеточным.
Во время дождя, когда ботридиум заливается водой,
внутри пузыря образуются многочисленные
зооспоры, выходящие из отверстия в верхней части.
Это напоминает извержение вулкана. Зооспоры
расселяются в воде, оседают на влажной почве и
развиваются в новые растения. Когда долго нет
осадков и почва пересыхает, содержимое надземной
части ботридиума образует апланоспоры или
переливается в ризоиды и там распадается на
цисты. При увлажнении цисты или прорастают в
новые особи, или формируют зооспоры.
В прудах, реках, озерах можно также встретить
планктонные желто-зеленые водоросли, имеющие
клетки разнообразной формы, и различные выросты
и украшения оболочек.
Диатомовые водоросли

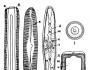
Диатомовая водоросль пиннулярия: вид со стороны пояска (1), со стороны шва (2), вскрытая клетка (3), поперечный срез через клетку (4), 5 – циклотелла; а – гипотека, б – эпитека, в – шов, г – узелок, д – хромопласт, е – пиреноиды, ж – цитоплазма, з – ядро, и – вакуоль
Отдел диатомовых включает более 10 000 видов водорослей, представленных исключительно одноклеточными или колониальными формами. Диатомеи распространены по всему земному шару и широко представлены в планктоне и бентосе морей и океанов, а также различных пресных водоемов, вплоть до горячих источников с температурой воды выше 50 оС. Эти водоросли можно встретить и в верховых болотах, на мхах, на камнях и скалах, в почве. Иногда они могут в массе развиваться на поверхности снега и льда, окрашивая их в бурый цвет. Ф.Нансен во время плавания на «Фраме» наблюдал появление на льду бурых пятен (скопления диатомей), под которыми лед быстро таял.
Большая часть клетки диатомовой водороли заполнена вакуолью, а цитоплазма располагается тонким слоем вдоль стенок. Но ядро обычно помещается в центре клетки, в особом цитоплазменном мостике. В цитоплазме располагаются пластинчатые или зернистые хлоропласты, в которых содержатся хлорофиллы а и с , а также каротиноиды и ксантофиллы, придающие водоросли желтую или желто-бурую окраску. Но после отмирания клетки диатомей зеленеют, т.к. хлорофилл разрушается позже, чем другие пигменты. Запасные питательные вещества диатомовых водорослей – масла, откладывающиеся в клетке в виде капель, а также волютин и хризоламинарин.
От других одноклеточных водорослей диатомовые резко отличаются тем, что снаружи к плазмалемме клетки плотно примыкает кремнеземовый панцирь, состоящий из двух половинок, надевающихся друг на друга, как крышка на коробку. Каждая такая половинка состоит из створки («донышка») и спаянного с ней пояскового кольца. Если смотреть на клетку диатомеи «в профиль», то видно, что пояски находят друг на друга.
Клетки различных диатомовых водорослей могут иметь (если смотреть со стороны донышка) либо радиальную, либо двустороннюю симметрию. Этот признак является систематическим – первых выделяют в класс Центрические, а вторых относят к классу Перистые, или Пеннатные.
У многих планктонных форм диатомей на панцире имеются также разнообразные выросты, увеличивающие площадь поверхности и способствующие парению в толще воды. Клетки ряда видов при помощи этих выростов могут соединяться друг с другом, образуя колонии.
Размножаются диатомеи вегетативным делением клеток, которое особенно интенсивно происходит весной. При делении масса протопласта увеличивается, и обе половинки панциря отодвигаются друг от друга. Ядро митотически делится, затем протопласт разделяется пополам в плоскости, параллельной створкам. После этого дочерние клетки одноклеточных диатомей расходятся, а у колониальных остаются соединенными своими створками.
Каждый новый протопласт наследует от материнской клетки половину панциря, а вторая образуется заново. Интересно, что у обеих дочерних клеток эта новая половинка будет меньшей («коробочкой»), в то время как старая в любом случае становится «крышечкой». В результате ряда последовательных делений размеры клеток в популяции прогрессивно уменьшаются.
Диатомовые водоросли – диплоидные организмы, гаплоидны у них только гаметы, образующиеся непосредственно перед половым процессом, который может происходить в форме изо-, гетеро- или оогамии. Образовавшаяся зигота сразу, без периода покоя, резко увеличивается в размерах и образует ауксоспору – «растущую спору» (от греч. «ауксо» – увеличиваю, расту). После завершения роста и созревания ауксоспоры в ней развивается новая клетка, у которой сначала образуется большая (внешняя), а затем меньшая (внутренняя) половина панциря. По размерам такие клетки значительно превышают родительские – таким образом, половой процесс приводит к восстановлению размеров клеток, измельчавших в результате вегетативного размножения.
Диатомовые водоросли играют большую роль в круговороте кремния в природе: как уже было сказано, в Мировом океане они ежегодно извлекают из воды и используют для построения своих панцирей до 150ґ109 т кремнезема, а отмирая, образуют особые геологические породы – диатомиты . Обладающие высокой пористостью и низким удельным весом, диатомиты широко используются в различных отраслях промышленности – из них изготавливают легкие кирпичи и применяют в качестве добавки к различным сортам цемента. Плитками из диатомита облицован купол собора святой Софии в Стамбуле. Очень широкое применение нашли диатомиты в качестве фильтрующего материала при производстве различных масел, жиров, в сахарной и химической промышленности.
Продолжение следует
9. Желтозеленые водоросли. Общая характеристика. Порядки.
Желто-зеленые водоросли могут иметь одноклеточные жгутиковые, коккоидные, нитчатые и сифональные формы. Хлоропласты дисковидной формы. Содержат пигменты хлорофилл a и с, каротиноиды. Запасные вещества - глюкан, жиры. Размножаются за счет деления клетки или половым путем (изогамия, оогамия).
Основные представители: ризохлорис (Rhizochloris), хлоротециум (Chlorothetium), трибонема (Tribonema), ботридиум (Botrydium).
10. Бурые водоросли. Строение, размножение, классы, основные представители, распространение, значение
К отделу бурых водорослей (около 1500 видов) относятся многочисленные макроскопические водоросли, общим внешним признаком которых служит желтовато-бурая окраска их слоевищ, обусловленная наличием желтых и бурых пигментов. Преимущественно дисковидные хлоропласты содержат хлорофиллы а и с, каротины и ксантофиллы. Пиреноиды очень мелкие. Запасные питательные вещества - ламинарин, маннит (сахароспирт), в небольших количествах - жир. Кроме обычных органелл в клетках присутствуют физоды, структуры, содержащие танины. Оболочки клеток снаружи ослизняющиеся - пектиновые, внутренний слой - целлюлоза.
Тип структуры таллома бурых водорослей преимущественно пластинчатый, нередко очень сложно организованный, редко разнонитчатый. Подавляющее большинство представителей этой группы – морские формы.
Слоевища бурых водорослей бывают размером от нескольких десятков микрометров до десятков метров (макроцистис – до 50 м). У высокоорганизованных форм талломы дифференцированы и напоминают цветковые растения. Некоторые крупные представители имеют воздушные пузыри. Все бурые водоросли растут прикрепленными к грунту или другим водорослям. Для прикрепления они формируют ризоиды или базальный диск. На поперечном разрезе крупных талломов бурых водорослей различаются зоны, занятые клетками разного строения, специализированными для выполнения отдельных функций.
Клетки одноядерные, одетые толстыми оболочками с крупными порами. Оболочка состоит из внутреннего целлюлозного и внешнего слоя, основу которого составляют белки, соединенные с альгиновой кислотой и ее солями. Альгулоза. Физоды. Хлоропласты обычно мелкие дисковидные, реже – лентовидные и пластинчатые. У ряда таксонов в хлоропластах имеется пиреноид. В хлоропластах тиллакоиды группируются в ламеллы по 3. Пигменты: хлорофилл a, c, каротиноиды. Запасные питательные вещества в клетках вне хлоропластов - ламинарин (полисахарид) и маннит (шестиатомный спирт), в небольших количествах липиды.
Размножение. У Б.В. встречаются все основные типы размножения – вегетативное, бесполое и половое. Вегетативное размножение происходит при случайном отделении ветвей от слоевища, у этих ветвей не образуются органы прикрепления и не формируются органы бесполого и полового размножения. Органы размножения образуются на них в случае, если их развитие к моменту отрыва слоевища от грунта уже началось. Бесполое размножение осуществляется зооспорами зооспорами (есть виды, размножающиеся моно- или тетраспорами). Строение зооспор. Зооспоры грушевидной формы имеют сбоку два неравных жгутика: передний - длинный, задний - короткий. Образование зооспор и гамет у бурых водорослей происходит во вместилищах двух основных типов: одногнездных и многогнездных. Мейоз происходит при формировании спор в одногнездных спорангиях. Половой процесс изогамный, гетерогамный и оогамный. Все бурые водоросли, за исключением фукусовых, имеют чередование поколений; на бесполом поколении (спорофите) в зооспорангиях (или тетраспорангиях) после редукционного деления образуются зооспоры (или тетраспоры); из них вырастают гаплоидные половые растения (гаметофиты) раздельнополые или обоеполые. После оплодотворения зигота без периода покоя развивается в новое бесполое диплоидное растение (спорофит). У фукусовых вся жизнь водоросли происходит в диплоидной фазе, гаплоидны лишь сперматозоиды и яйцеклетки, перед образованием которых происходит редукционное деление.
В зависимости от характера чередования поколений и смены ядерных фаз бурые водоросли делят на 3 класса.
Класс Изогенератные (Isogeneratae)
У большинства водорослей этого класса спорофит и гаметофит одинаковы по форме и величине или не очень резко отличаются по величине.Половой процесс изо-, гетеро- или оогамный. Широко распространены в морях представители рода эктокарпус (Ectocarpus); особенно много их в холодных морях. Произрастают в литорали и сублиторали. Участвуют в обрастании судов, буйков. Они имеют вид маленьких кустиков или дерновинок, состоящих из обильно ветвящихся однорядных нитей, оканчивающихся нередко бесцветными многоклеточными волосками. Рост нитей интеркалярный. Сбоку на ветвях образуются одноклеточные зооспорангии. В них происходит редукционное деление и еще ряд делений клеточного ядра и формируется
много зооспор, которые выходят из зооспорангия, недолго поплавав, прорастают в такое же по внешности, но гаплоидное растение. На нем, на коротких боковых веточках образуются многоклеточные гаметангии, в которых развиваются одинаковые по форме и величине, но отличающиеся по поведению (+ и -) гаметы. Зигота без периода покоя прорастает в новую диплоидную нить с зооспорангием. Для эктокарпуса характерна изоморфная смена поколений. Половой процесс - изогамия.
Кутлярия (Cutleria) - распространена у европейских берегов Атлантического океана и в Средиземном море. Она имеет вид дихотонически разветвленных многослойных лент до 20 см длины, оканчивающихся волосками. Внешне кутлериевые отличаются от других водорослей по наличию бахромы из нежных волосков на вершине или по краю слоевища. У основания волосков на одних экземплярах развиваются крупноклеточные макрогаметангии, дающие двужгутиковые макрогаметы, на других экземплярах - развиваются мелкоклеточные микрогаметангии, дающие микрогаметы. После оплодотворения зигота без периода покоя развивается в новое диплоидное растение - спорофит, имеющее форму многослойной пластинки или корочки, плотно прижатой к субстрату. Она настолько не похожа на гаметофит, что была описана под новым родовым названием аглаозонии (Aglaozonia). На верхней стороне ее развиваются одноклеточные зооспорангии, в которых после редукционного деления формируются зооспоры. Зооспоры прорастают в лентовидный кустистый гаметофит. Род кутлярия пользуется наибольшей известностью. Она служит примером гетероморфной смены поколения, половой процесс ее - гетерогамия.
Диктиота (Dictyota) произрастает преимущественно в тропических и субтропических морях, встречается и в Черном море. Для нее характерно вильчато-разветвленное слоевище с расположенными в одной плоскости ветвями. Высота растения около 20 см, ширина ветвей 4-8 мм. Диктиота имеет изоморфное чередование поколений. На спорофите из поверхностных клеток развиваются крупные шаровидные тетраспорангии, в которых после редукционного деления образуется 4 гаплоидные неподвижные голые тетраспоры. Из тетраспор развиваются мужские (с многочисленными антеридиями, дающими до 30000 сперматозоидов) и женские (с оогониями, продуцирующими по одной яйцеклетке) гаметофиты. Зрелые яйцеклетки выпадают из оогониев. После оплодотворения зигота развивается в новый спорофит. Диктиота - пример развития водоросли с изоморфной сменой поколений, у которой половой процесс - оогамия.
Класс Гетерогенератные (Heterogeneratae)
В цикле развития водорослей этого класса чередуются макроскопический (у многих мощно развитый) спорофит и микроскопический мелкий гаметофит, который нередко по аналогии с папоротниками называется заростком. Половой процесс изогамный или оогамный. Спорофиты разных родов резко различаются и представляют собой самые крупные низшие растения достигающие в длину 60-100 м со значительным морфологическим расчленением и сложным анатомическим строением. Спорофиты расчленяются на листовую пластинку, черешок и ризоиды или базальный диск. В месте перехода пластинки в черешок - интеркалярная меристема. Спорофит многолетний, за исключением пластинки, которая ежегодно отмирает и заменяется новой. В меристодерме пластинки перед ее разрушением образуются одноклеточные зооспорангии. В каждом зооспорангии после предварительного редукционного деления образуется от 16 до 64 (иногда 128) зооспор. На одном экземпляре водоросли формируются миллиарды зооспор. После недолгого периода движения зооспоры прорастают в микроскопически мелкие нитевидные мужские и женские гаметофиты (заростки). В антеридиях образуется по одному сперматозоиду, в оогониях - по одной яйцеклетке. После оплодотворения из зиготы развивается постепенно новый мощный спорофит.
Итак, для гетерогенаратных водорослей характерна гетероморфная смена поколений. Половой процесс - оогамия.
Ламинария (Laminaria) , виды рода широко распространены в северных морях. Таллом расчленен на листовую пластинку, ствол и ризоиды. Встречается по вертикали от поверхности моря до глубины 200 м.
Макроцистис (Macrocystis) , растущий на глубине 10-20 м, имеет ствол многократно разветвленный до 50-60 м длиной. Перед каждой пластинкой ветвь обычно расширена в грушевидный воздушный пузырь. Благодаря воздушным пузырям и длинным ветвям основная часть слоевища макроцистиса плавает у поверхности моря. Его добывают во всем мире и перерабатывают для получения альгинатов и других химических продуктов.
Нереоцистис (Nereocystis) представлен длинным стволом (черешком) до 15-25 м, который постепенно расширяется кверху, несет на конце пузырь диаметром 12-20 см, от которого отходят узкие пластинки (до 9 м).
Всего таких пластин 24-40. Растет нереоцистис на глубине до 20 м, при этом верхняя часть ствола с пузырем плавает у поверхности.
Жизненные циклы вышеназванных представителей схожи. Зооспорангии формируются на листовых пластинках.
Класс Циклоспоровые (Cyclosporeae)
К этому классу относятся бурые водоросли, не имеющие чередования поколений, а лишь смену ядерных фаз: вся водоросль диплоидна, гаплоидны лишь гаметы. Бесполого размножения нет. Класс содержит всего один порядок - фукусовые (Fucales).
Фукусы имеют кожистые, оливко- или желто-бурые, дихотонически разветвленные ремневидные талломы длиной 0,5-1 м, 1-5 см ширины; у некоторых имеются вздутия, наполненные воздухом. Рост верхушечный.
Ко времени полового размножения на концах ветвей образуются бобовидные желтоватые вздутия - рецептакулы, на которых располагаются половые органы. Гаметофиты развиваются на рецептакулах в виде слоя, выстилающего углубления (концептакулы или скафидии). Гаметангии формируются в углублениях на слое клеток, который возникает из одной клетки, именуемой инициальной клеткой концептакула или проспорой. Развивающийся из проспоры выстилающий слой концептакула - гаметофит. Гаметофиты фукусовых образуют вертикальные разветвленные и простые одноядерные многоклеточные нити, часть из них несет гаметангии, а остальные служат парафизами.
Оогонии у фукусовых сидят прямо на поверхности гаметофита. Антеридии образуются на концах разветвленных веточек. Развивающиеся оогонии (8 яйцеклеток) и антеридии (64 сперматозоида) слизью вытал-
киваются из концентакула наружу, здесь из них освобождаются половые клетки. Оплодотворение происходит в окружающей воде. Оплодотворенная яйцеклетка без периода покоя прорастает в новое растение.
Род фукус (Fucus) распространен в северных морях, это основной обитатель береговой зоны.
Саргассум (Sargassum) - «листья» пластинчатые или шиловидные. Ствол короткий, от верхней части его отходят длинные ветви, несущие рецептакулы, эти ветви ежегодно отмирают. Представители распространены в основном в тропических и субтропических морях. Саргассум используют для получения альгинатов, отдельные виды с нежесткими листьями используют в пищу. В Саргассовом море на площади 4,4 млн. кв. км плавают два вида саргассума, которые размножаются вегетативно и не имеют органов прикрепления.
Изоморфный или гетероморфный варианты гаплоидно-диплоидного жизненного цикла или изоморфная и гетероморфная смены генераций. Жизненные циклы родов Ectocarpus, Laminaria, Fucus, концептакулы, рецептакулы или скафидии.
Распространение и экология Б.В. Почти исключительно морские растения. Распространены они повсюду от Арктики до Антарктики. Но наиболее крупные виды растут преимущественно в умеренных и приполярных широтах. Представители отдела растут в основном на мелководье, преобладают на каменистых прибрежных субстратах в холодных районах земного шара. но встречаются и на глубинах до 200 м. Виды, имеющие воздушные пузыри, при отрыве от грунта становятся свободно плавающими и иногда (напр. в Саргассовом море) образуют большие скопления на поверхности воды. Наиболее мощные заросли Б.В. образуют в водах, богатых биогенными веществами. Многие крупные водоросли широко используются на корм скоту и для удобрений полей (богаты калием). Некоторые виды ламинарии используют в пищу, помимо пищевого, они имеют и диетическое значение, т.к. богаты йодом и полезны при атеросклерозе. Раньше из ламинарий добывали йод. В значительных количествах используют бурые водоросли для получения альгина, обладающего большой склеивающей способностью и применяемого при производстве бумаги, картона, изготовлении типографских красок. Водоросли собирают особыми жатвенными машинами, а также используют выброшенные на берег.
План:
-
Введение
- 1
Строение клетки
- 1.1 Жгутики
- 1.2 Хлоропласт
- 1.3 Клеточная стенка
- 1.4 Другие структуры
- 1.5 Ядро
- 2 Размножение
- 3 Экология
- 4 Значение
- 5 Филогения
- 6 Разнообразие и систематика Примечания
Литература
Введение
Жёлто-зелёные водоросли (лат. Xanthophyceae , или Xanthophyta ), или Разножгутиковые водоросли (лат. Heterocontae ), или Трибофициевые (лат. Tribophyceae ) - класс низших растений, включающий в себя водоросли, хлоропласты которых окрашены в жёлто-зелёный или жёлтый цвет. Представители - одноклеточные, колониальные и многоклеточные, преимущественно пресноводные организмы. Аналогично золотистым водорослям, в основу деления жёлто-зелёных на классы положено разнообразие морфологической организации таллома. Класс назван по типовому роду Tribonema (от греч. tribon - опытный, искусный, nema - нить).
1. Строение клетки
1.1. Жгутики
У монадных представителей (у зооспор и гамет) имеются два неравных по длине и морфологии жгутика: на главном жгутике расположены перистые мерцательные волоски, боковой жгутик - бичевидный. Исключением являются синзооспоры Vaucheria , у которых по поверхности расположены многочисленные пары немного различающихся по длине гладких жгутиков. Жгутики прикрепляются на клетке субапикально (у сперматозоида Vaucheria прикрепление латеральное). Мастигонемы синтезируются в цистернах эндоплазматической сети. Короткий жгутик заканчивается акронемой.
Базальные тела жгутиков трибофициевых типичного строения, расположены под прямым углом друг к другу. Корешковая система представлена поперечноисчерченным корешком - ризопластом и тремя микротрубочковыми корешками, каждый из которых состоит из 3-4 микротрубочек.
1.2. Хлоропласт
Хлоропласт имеет типичное для охрофитов строение. Обычно в клетке присутствует несколько зелёных или жёлто-зелёных дисковидных пластид. Их окраска связана с отстуствием фукоксантина, отвечающего за золотистый и коричневый цвет у других охрофитовых. Из каротиноидов у трибофициевых присутствует α- и β-каротины (преобладают), вошериаксантин, диатоксантин, диадиноксантин, гетероксантин, лютеин, виолаксантин, неоксантини др. Хлорофиллы - a и c . В клетках трибофициевых кроме дисковидных встречаются пластиды и других форм: пластинчатые, корытовидные, лентовидные, чашевидные, звёздчатые и др. У немногих видов обнаружены пиреноиды полупоргужённого типа. Глазок состоит из ряда липидных глобул, расположен на переднем конце тела в хлоропласте, ориентирован на базальное вздутие жгутика.
1.3. Клеточная стенка
У видов с амебоидной, монадной и пальмеллоидной организацией клеточная стенка отсутствует, они покрыты только цитоплазматической мембраной и могут легко менять форму. Иногда «голые» клетки находятся внутри домиков, стенки которых могут быть окрашены в бурый цвет солями марганца и железа. У подавляющего большинства трибофициевых имеется клеточная стенка цельная или состоящая из двух частей. В её составе, изученном у Tribonema и Vaucheria , преобладает целлюлоза и содержатся полисахариды, состоящие преимущественно из глюкозы и уроновых кислот. У молодых клеток оболочка тонкая, с возрастом она утолщается. В ней могут откладываться соли железа, соединения которого окрашивают её в различные оттенки коричневого и красного тонов. Чаще в клеточной стенке присутствует кремнезём, придавая ей твёрдость и блеск. Она может инкрустироваться также известью и быть различным образом скульптурирована (шипики, ячейки, бородавки, щетинки, зубчики и т. д.). У прикреплённых форм может образовываться вырост оболочки - ножка с прикрепительной подошвой.
У нитчатых водорослей с двухстворчатыми оболочками при распаде нитей клеточные оболочки разваливаются на Н-образные фрагменты, представляющие собой плотно соединённые половинки оболочек двух соседних клеток. При росте нитей Н-образный фрагмент клеточной стенки двух соседних дочерних клеток встраивается между двумя половинками оболочки материнской клетки. В результате этого каждая из дочерних клеток наполовину покрыта старой оболочкой материнской клетки и наполовину - новообразованной оболочкой.
1.4. Другие структуры
Сократительные вакуоли имеются у подвижных представителей. Обычно их 1-2 на клетку, иногда больше. Аппарат Гольджи своеобразного строения. Диктиосомы мелкие, содержат 3-7 цистерн.
Запасные питательные вещества - масла, у некоторых - волютин, хризоламинарин и лейкозин.
1.5. Ядро
Ядро одно, реже ядер много, у ценотических представителей клетки всегда многоядерные. Детали митоза подробно изучены только у Vaucheria . Митоз у неё закрытый, с центриолями, расположенными на полюсах вне ядра. Кинетохоры не обнаружены. Во время анафазы сильно удлиняются межполюсные микротрубочки веретена, что приводит к значительному удалению дочерних ядер друг от друга. Ядерная мембрана сохраняется, поэтому в телофазе дочерние ядра имеют вид гантели. Полагают, что такой митоз не является типичным для всей группы трибофициевых.
2. Размножение
У большинства жёлто-зелёных известно вегетативное и бесполое размножение. Вегетативное размножение осуществляется делением клеток пополам, распадом колоний и многоклеточных слоевищ на части. При бесполом размножении могут формироваться амебоиды, зооспоры, синзооспоры, гемизооспоры, гемиавтоспоры, автоспоры, апланоспоры. Зооспоры «голые» и обычно имеют грушевидную форму, два жгутика. Половой процесс (изо-, гетеро- и оогамный) описан у немногих представителей.
При наступлении неблагоприятных условий наблюдается образование цист. Цисты (статоспоры) эндогенные, одноядерные, реже многоядерные. Их стенка часто содержит кремнезём и состоит из двух неравных, или реже - равных частей.
3. Экология
Трибофициевые встречаются на всех континентах, в том числе в Антарктиде. Они обитают преимущественно в пресных водах умеренных широт, обычны также в почве, реже встречаются в наземных, солоноватоводных и морских местообитаниях. Населяют как чистые, так и загрязнённые воды, с различным значением pH, но редко встречаются в обилии. Значительно разнообразнее и обильнее трибофициевые водоросли представлены в почвах, где, развиваясь в массе, могут вызвать «цветение» её поверхности. Аэрофитные представители встречаются на стволах деревьев, скалах, стенах домов, иногда вызывая их позеленение. Часто обитают в скоплениях нитчатых водорослей и водных высших растений по берегам рек, прудов, озёр и водохранилищ.
Жёлто-зелёные водоросли входят в различные экологические группы - планктон, реже в перифитон и бентос. Подавляющее большинство из них - свободноживущие формы, но встречаются и внутриклеточные симбионты - зооксантеллы в клетках простейших. Интересный внутриклеточный симбиоз образуют хлоропласты морской водоросли V. litorea с моллюском Elysia chlorotica . На протяжении 9 месяцев этот моллюск способен в культуре к фотоавтотрофной фиксации углекислого газа. Это самый длительный по времени симбиоз такого типа, когда симбиотическая пластида находится в непосредственном контакте с цитоплазмой животного. В природе личинки моллюска питаются нитями Vaucheria . В результате фагоцитоза хлоропласты водоросли попадают в цитоплазму эпителиальных клеток моллюска. В ходе этого процесса оболочка хлоропласта становится трёхслойной, теряется наружная мембрана хлоропластной эндоплазматической сети. Такое явление может служить доказательством того, что в процессе эволюции в результате вторичного симбиогенеза за счёт потери мембран могли возникать хлоропласты с тремя мембранами.
4. Значение
Трибофициевые водоросли - продуценты кислорода и органических веществ, входят в состав трофических цепей. Они участвуют в самоочищении загрязнённых вод и почв, образовании илов и сапропелей, в процессе накопления органических веществ в почве, влияя на её плодородие. Их хозяйственное значение сводится к использованию как показательных организмов при определении состояния загрязнения вод; они входят в состав комплекса микроорганизмов, применяемого для очистки сточных вод.
5. Филогения
В конце XIX - начале XX вв. различные роды трибофициевых относили к отделу зелёных водорослей, что было связано в первую очередь с окраской и морфологическим сходством талломов. Но уже А. Пашер включил эту группу в один эволюционный ряд с золотистыми и диатомовыми водорослями. Эта точка зрения в дальнейшем нашла подстверждение в исследованиях на цитологическом, биохимическом и молекулярном уровне. В настоящее время трибофициевые рассматриваются как класс внутри отдела охрофитовых. Из трибофициевых были выделены эвстигматофициевые в ранге такого же класса, но, как оказалось, в эволюционном плане они далеко стоят друг от друга. В филогенетических деревьях, построенных на анализе нуклеотидных последовательностей ряда генов, трибофициевые среди охрофитовых стоят значительно ближе к бурым водорослям, чем к золотистым, диатомовым, синуровым и эвстигматофициевым.
6. Разнообразие и систематика
Описано около 90 родов и более 600 видов, которые группируются в 6-7 порядков (H. Ettl, 1978). В основу выделения порядков положены тип дифференциации таллома и особенности жизненного цикла. Количество порядков зависит от точки зрения на ценотические трибофициевые водоросли: относят ли их к одному или двум порядкам.
Примечания
- По данным EOL. См. карточку.
Литература
- Жизнь растений. В 6-ти т. / Гл. ред. Ал. А. Фёдоров. - М .: Просвещение, 1977. - Т. 3. Водоросли. Лишайники . Под ред. М. М. Голлербаха. - 487 с. - 300000 экз .
- «Курс низших растений», под ред. М. В. Горленко - М.: «Высшая школа», 1981
- В. А. Крыжановский, Г. Л. Билич «Биология. Полный курс. В 3-х тт. Том 2. Ботаника.» - М.: ООО "Издательский дом «ОНИКС 21 век», 2002
- «Малый практикум по низшим растениям» - М.: «Высшая школа», 1976
- Н. П. Горбунова «Альгология», М.: «Высшая школа», 1991
К Желтозеленым относятся водоросли, хлоропласты которых окрашены в светло- или темно-желтый цвет, очень редко зеленый и лишь иногда в голубой. Окраска талломов определяется наличием в хлоропластах клеток следующих пигментов – хлорофиллов а и с , β -каротина и ксантофиллов. Преобладание последних и обусловливает своеобразие окраски желтозеленых водорослей. Кроме того, в клетках в качестве основного продукта ассимиляции накапливаются парамилон, капли масла и лишь у некоторых видов, кроме того, глыбки лейкозина и волютина. Крахмал у желтозеленых водорослей не образуется. Отличительной особенностью желтозеленых является наличие у вегетативных клеток монадной структуры и у зооспор двух неравных жгутиков. В состав клеточной стенки входят целлюлоза, глюкоза и уроновые кислоты. Клеточная стенка часто состоит из двух частей.
Размножение вегетативное, бесполое и половое.
Широко распространены в пресных водах. Редко встречаются в морских, солоноватых водах и в почве.
Ранее класс Желтозеленые водоросли назывался Трибофициевые водоросли по типовому роду Трибонема (от греч. tribon – искусный, хитрый и nema – нить). Известно около 450 видов.
Желтозеленые характеризуются значительным морфологическим разнообразием. Среди многочисленных представителей этого отдела обнаруживаются почти все основные типы структуры тела: амебоидная, монадная, пальмеллоидная, коккоидная, нитчатая, разнонитчатая, пластинчатая и сифональная (рис. 44– 46). Слоевище одноклеточное,
Рис. 44. Внешний вид желтозеленых водорослей: 1, 2 – Харациопсис , 3 – Центритрактус , 4 – Офиоцитиум
колониальное, многоклеточное и неклеточное. Клеточная оболочка плотная, пектиновая и целлюлозная, состоящая из плотно налегающих друг на друга частей или из двух створок. В оболочке откладывается кремнезем или известь. Преимущественно неподвижные формы. Среди одноклеточных видов имеются подвижные формы, лишенные плотной оболочки и снабженные жгутиками, лобоподиями и ризоподиями.

Рис. 45. Внешний вид ксантококковых желтозеленых водорослей: 1–3 – Ботридиопсис , 4 – Тетраэдриелла , 5 – Псевдостаураструм , 6 – Гониохлорис , 7, 8 – Бумиллериопсис
Большинство желтозеленых – неподвижные организмы. У подвижных особей движение может осуществляться с помощью жгутиков либо ризоподий. Клетки различной формы: шаровидные, веретеновидные, эллипсоидные, цилиндрические, тетраэдрические, серповидные, грушевидные, яйцевидные. Талломы размерами от 0.5– 1.5 мкм (Хлориделла ) до нескольких миллиметров в диаметре (Ботридиопсис ) (рис. 45, 1– 3) и до десятков сантиметров в длину (Вошерия ) (рис. 46, 3).

Рис. 46. Внешний вид желтозеленых водорослей: 1 – Трибонема , 2 – Гетеропедия , 3 – Вошерия , часть нити с оогонием и антеридием
Большинство видов желтозеленых являются фототрофами, но встречается и голозойное питание путем заглатывания бактерий и мелких водорослей. Желтозеленые водоросли широко распространены в пресных водах. Они также обычны в почве, реже встречаются в морских и солоноватоводных водоемах. Класс включает аэробионтные, планктонные, донные и перифитонные формы. Эпифиты, эпизоиты, а также внутриклеточные симбионты в клетках простейших.
Независимо от внешней структуры, внутреннее строение клетки желтозеленых водорослей однотипно. В протопласте наблюдается обычно несколько желто-зеленых хлоропластов, имеющих дисковидную, корытовидную, пластинчатую, реже лентовидную, звездчатую или чашевидную форму с цельными или лопастными краями. Окраска связана с отсутствием фукоксантина, отвечающего за золотистый и коричневый цвет у других охрофитовых. Из других пигментов у них имеются β -каротин, вошериаксантин, диатоксантин, диадиноксантин, гетероксантин. У подвижных форм на переднем конце хлоропласта обычно расположен красный глазок, или стигма. У немногих видов имеются пиреноиды полупогруженного типа. Ядро в клетке обычно одно, небольших размеров, но имеются виды и с многоядерными клетками. У некоторых видов в передней части клетки имеется одна или две сократительные (пульсирующие) вакуоли.
У монадных представителей и у подвижных стадий (зооспор и гамет) имеются два неравных жгутика. Исключением являются синзооспоры Вошерии , у которых по поверхности расположены многочисленные пары немного различающихся по длине гладких жгутиков. Короткий жгутик заканчивается акронемой. Жгутики прикрепляются на клетке субапикально. У сперматозоида Вошерии прикрепление латеральное.
У видов с амебоидной, монадной и пальмеллоидной организацией клеточная стенка отсутствует, они покрыты только цитоплазматической мембраной и могут легко менять форму. Иногда «голые» клетки находятся внутри домиков, стенки которых могут быть окрашены в бурый цвет солями марганца и железа. У подавляющего большинства форм имеется клеточная стенка, состоящая из двух частей. В составе клеточной стенки преобладает целлюлоза, а также содержатся полисахариды, состоящие в основном из глюкозы и уроновых кислот. У молодых клеток оболочка тонкая, с возрастом она утолщается. В ней могут откладываться соли железа, соединения которого окрашивают ее в различные оттенки коричневого и красного тонов. Чаще в клеточной стенке присутствует кремнезем, придавая ей твердость и блеск. Она может инкрустироваться также известью и быть различным образом скульптурирована (шипики, ячейки, бородавки, щетинки, зубчики и т.д.) У прикрепленных форм может образовываться вырост оболочки – ножка с прикрепительной подошвой.
У нитчатых форм желтозеленых водорослей с двухстворчатыми оболочками при распаде нитей клеточные оболочки разваливаются на Н-образные фрагменты. Эти фрагменты представляют собой плотно соединенные половинки оболочек двух соседних клеток (рис. 47). При росте нитей Н-образный фрагмент клеточной стенки двух соседних дочерних клеток встраивается между двумя половинками оболочки материнской клетки. В результате каждая из дочерних клеток наполовину покрыта старой оболочкой материнской клетки и наполовину – новообразованной оболочкой.

Рис. 47. Схема образования поперечной перегородки между двумя дочерними клетками у нитчатых желтозеленых водорослей (по: А.А. Масюк, 1993): А – фрагмент нити; Б – закладка пояскового кольца и образование поперечной перегородки между двумя клетками; В – слоистость двустворчатых клеточных оболочек; Г – распад оболочки на Н-образные участки
Сократительные вакуоли имеются у подвижных представителей. Обычно их 1-2 на клетку. Аппарат Гольджи своеобразного строения. Диктиосомы мелкие, содержат 3-7 цистерн. Ядро одно, реже их много; у ценотических видов клетки всегда многоядерные.
Размножение . Для большинства видов желтозеленых водорослей характерно вегетативное и бесполое размножение.
Вегетативное размножение осуществляется различными способами: делением клеток пополам, распадом колоний и многоклеточных слоевищ на части. У Вошерии образуются специальные выводковые почки.
При бесполом размножении могут формироваться разнообразные споры: амебоиды, зооспоры, синзооспоры, автоспоры, гемизооспоры, гемиавтоспоры, апланоспоры. Зооспоры «голые» и обычно грушевидной формы.
Половой процесс – изогамия, гетерогамия и оогамия – описан у немногих представителей. У Трибонемы гаметы сходны по размерам, но различаются поведением – это изогамия. У Вошерии наблюдается оогамия: на нитях образуются вместилища женских гамет – оогонии и мужских – антеридии.
В неблагоприятных условиях наблюдается образование цист. Цисты (статоспоры) эндогенные, одноядерные, реже многоядерные. Их стенка часто содержит кремнезем и состоит из двух неравных или, реже, равных частей.
Систематика.
В конце XIX – начале XX вв. различные роды желтозеленых относили к отделу зеленых водорослей, что было связано в первую очередь с окраской и морфологическим сходством талломов. В настоящее время желтозеленые рассматриваются как класс внутри отдела охрофитовых.
Известно около 450 современных видов класса Желтозеленые водоросли, которые группируются в четыре порядка: Ботридиевые, Мишококковые, Трибонемовые и Вошериевые. В основу выделения порядков положены тип дифференциации таллома и особенности жизненного цикла.
Порядок Ботридиевые – Botrydiales . К порядку относятся виды с сифональным типом дифференциации таллома, у которых отсутствует оогамный половой процесс.
Род Ботридиум обитает на почве и имеет вид зеленых пузырей величиной несколько миллиметров, прикрепленных с помощью бесцветных ризоидов. Таллом сифональный, содержит многочисленные ядра и пластиды. Оболочка многослойная, на ней может откладываться известь. Размножение бесполое с помощью двухжгутиковых зооспор, при этом все содержимое пузыря распадается на одноядерные фрагменты. При недостатке влаги размножается с помощью апланоспор или формирует толстостенные цисты. В одних случаях все содержимое пузыря идет на образование одной крупной цисты. В других случаях цисты формируются в ризоидах, куда вначале переходит содержимое пузыря. Цисты прорастают или непосредственно в новый таллом, или образуют зооспоры. Половой процесс – изо- и гетерогамия. Зигота прорастает немедленно, без периода покоя. Обычные и широко распространенные в наземных местообитаниях виды, встречаются по берегам ручьев, прудов или на лишенных растительности почвах.
Порядок Мисхококковые –Mishococcales . Одноклеточные или колониальные представители с коккоидным типом дифференциации таллома.
Род Харациопсис включает одноклеточные прикрепленные формы. При размножении формирует зооспоры, апланоспоры и толстостенные цисты (рис. 44, 1-2).
Род Офиоцитиум (рис. 44, 4) имеет удлиненные цилиндрические клетки, которые могут быть прямыми, согнутыми или спирально закрученными, а на конце могут нести шип. Клеточная стенка состоит из двух неравных частей, из них большая часть участвует в росте клетки, меньшая часть постоянна и имеет форму крышечки. Одноклеточные и колониальные виды, свободноживущие или прикрепленные к субстрату с помощью маленькой ножки. Размножаются зооспорами и апланоспорами, встречаются цисты. Обитают в пресных водах.
Род Мисхококкус образует древовидные прикрепленные колонии. Ветвление дихотомическое и тетрахотомическое. Клетки расположены по 2 или 4 на верхушках слизистых ветвей колонии. Клетки шаровидные, до овальных, с тонкой или толстой клеточной стенкой. Иногда клеточная стенка блестящая и коричневая из-за пропитки ее солями железа. Молодые одноклеточные организмы со слизистым дисковидным основанием, которое служит точкой опоры при прикреплении. После выхода спор протопласт материнской клетки превращается в студень и вытягивается, длина становится в 6 раз больше ширины, и таким образом возникает цилиндрическая ножка. Пустая клеточная стенка материнской клетки всегда становится основанием ножки. Бесполое размножение зооспорами и автоспорами. Автоспоры прикрепляются к верхнему краю слизистой ножки. Последующие клеточные деления повторяют процесс и производят древовидную колонию. Половой процесс – изогамия. Обитают в небольших пресных водоемах как эпифиты нитчатых водорослей. Известны в центральной Европе и Азии.
Порядок Трибонемовые – Tribonematales . Представители имеют нитчатый, разнонитчатый, ложнотканевый и тканевый тип дифференциации таллома. Клеточные стенки либо с Н-образными перекрывающимися частями, либо цельные.
Род Трибонема – неветвящиеся нити (рис. 46, 1). Клетки по форме цилиндрические или бочонкообразные. Клеточная стенка состоит из двух половинок, которые находят краями друг на друга в середине клетки. Оболочки часто слоистые. Фрагменты нитей всегда оканчиваются пустыми половинками Н-образных фрагментов оболочки, имеющих вид вилочки. В клетках несколько желтовато-зеленых пластид, пиреноидов нет. Размножение вегетативное (фрагментацией нитей), бесполое (зооспорами и апланоспорами) и половое (изогамия), причем апланоспоры образуются чаще, чем зооспоры. Могут формировать акинеты. Обитают в пресных водах, где особенно обильно развиваются в холодное время года.
Порядок Вошериевые – Vauсheriales . Все представители имеют сифональный таллом, оогамный половой процесс и синзооспоры.
Род Вошерия (рис. 46, 3) имеет слоевище неклеточного строения; ее таллом достигает в длину нескольких сантиметров, прикрепляется к субстрату с помощью бесцветного ризоида. В нитях отсутствуют перегородки, большая часть таллома занята вакуолью, а по периферии в цитоплазме расположены многочисленные ядра и пластиды. Нити с апикальным ростом и редким боковым ветвлением. Перегородки образуются при повреждении таллома и для отделения органов размножения. Бесполое размножение осуществляется апланоспорами, синзооспорами, акинетами. Синзооспоры формируются по одной в зооспорангии, который отделяется от вегетативных клеток перегородкой на конце нити. Зооспоры многоядерные и многожгутиковые. Половой процесс – оогамия. Зигота покрывается толстой оболочкой и после периода покоя прорастает в новый таллом.
Виды Вошерии широко распространены в пресных, солоноватых и морских водах, а также в наземных местообитаниях. Они встречаются на всех континентах, включая Антарктиду. Формируют травянисто-зеленые или темно-зеленые перепутанные массы – так называемые маты, гладкие, стелющиеся или подушковидные. Водные, полуводные, наземные формы. Обитают в разнообразных местообитаниях: морях, эстуариях, лиманах, соленых маршах, мангровых зарослях, ручьях, каналах, озерах, прудах, на пахотных землях и болотах.
Значение гетероконтных водорослей
Из отдела Гетероконтные водоросли наибольшее значение для природных экосистем и для человека имеют бурые водоросли.
Бурые водоросли – основной источник органического вещества в прибрежной зоне морей. Их биомасса в морях умеренных и приполярных зон может достигать нескольких десятков килограммов на квадратный метр. Заросли бурых водорослей создают условия для питания и размножения множеству прибрежных животных и других водорослей. Чарльз Дарвин, наблюдавший у берегов Южной Америки заросли бурой водоросли Макроцистиса , писал: «Эти огромные подводные леса Южного полушария я могу сравнить только с наземными лесами тропических областей. И все-таки, если бы в какой-нибудь стране уничтожить лес, то не думаю, чтобы при этом погибло хотя бы приблизительно такое количество видов животных, как с уничтожением этой водоросли».
Заросли бурых водорослей служат местом питания, укрытия и размножения многих животных. Образно говоря, бурые водоросли предоставляют другим водным организмам «стол, кров и ясли».
Широко используются бурые водоросли и человеком. Они богаты йодом и другими микроэлементами. Народы Юго-Восточной Азии традиционно используют их в пищу , особенно представителей порядка ламинариевых, из которых готовят множество самых разнообразных блюд. Кормовая мука , приготавливаемая из бурых водорослей, повышает продуктивность скота; при этом в яйцах и молоке увеличивается содержание йода.
Из бурых водорослей получают альгинаты – соли альгиновой кислоты. Альгинаты очень широко применяются в различных областях промышленности. Это нетоксичные соединения, обладающие коллоидными свойствами, поэтому они широко используются в пищевой и фармацевтической промышленности. Альгиновая кислота и ее соли способны к 200–300-кратному поглощению воды, образуя гели, для которых характерна высокая кислотоустойчивость. В пищевой промышленности они используются преимущественно в качестве эмульгаторов, стабилизаторов, желирующих и влагоудерживающих компонентов. Например, сухой порошковый альгинат натрия используют в производстве порошкообразных и брикетированных растворимых продуктов (кофе, чай, сухое молоко, кисели и др.) для их быстрого растворения. Водные растворы альгинатов используют для замораживания мясных и рыбных продуктов. В мире в пищевую промышленность идет до 30% всего объема получаемых альгинатов.
В текстильной и целлюлозно-бумажной промышленности альгинаты используют для загущения красок и усиления прочности их связи с основой. Пропитка тканей некоторыми альгинатами придает им защитные свойства: водонепроницаемость, кислотоустойчивость и увеличивает механическую прочность. Ряд солей альгиновых кислот используют для получения искусственного шелка. Во времена Второй мировой войны в США и Англии из альгиновой кислоты и ее солей производилось большое количество маскировочной ткани и сетей для жилых и промышленных зданий.
Альгинаты применяют в металлургии: в литейном производстве они улучшают качество формовочной земли. Соли альгиновой кислоты применяют в производстве электродов для электросварки, позволяющих получать более высококачественные сварные швы. Альгинаты также используют при производстве пластмасс, синтетических волокон, лакокрасочных покрытий и строительных материалов, устойчивых к атмосферным воздействиям. Их применяют при изготовлении высококачественных смазочных материалов для машин. В радиоэлектронике альгинаты выполняют роль связующего агента при изготовлении высококачественных ферритов.
Наибольшее применение находит водорастворимый альгинат натрия, способный к образованию вязких растворов. Его широко используют для стабилизации разнообразных растворов и суспензий. Добавление небольшого количества альгината натрия в пищевые продукты – консервы, мороженое – повышает их качество. Он также используется для изготовления декоративной косметики, кремов и масок в парфюмерной промышленности.
В фармацевтической промышленности альгинаты используются для покрытия таблеток, пилюль, в качестве компонентных основ для различных мазей и паст, как гели-носители лекарственных препаратов, в производстве растворимых хирургических нитей. В медицине альгинат кальция применяют в качестве кровеостанавливающего средства и как сорбент, выводящий радионуклиды (например, стронций). Ежегодное производство альгинатов в мире превышает 20 тысяч тонн.
Другое важное вещество, получаемое из бурых водорослей, – шестиатомный спирт маннит. Маннит используют как заменитель сахара для больных диабетом. Кроме того, он может быть использован в качестве плазмозаменителя при консервации крови. Его применяют для изготовления таблеток в фармацевтической промышленности. Маннит также используют в производстве синтетических смол, красок, бумаги, взрывчатых веществ, при выделке кож.
Фукоиданы, получаемые из бурых водорослей – эффективные антикоагулянты, даже более активные, чем гепарин. Перспективным считается использование их для получения противоопухолевых препаратов и антивирусных соединений. Ведь даже в самых низких концентрациях фукоиданы могут ингибировать прикрепление вирусов к поверхности клеток. Фукоиданы также обладают способностью образовывать исключительно прочные и вязкие слизи, что находит применение в получении стабильных эмульсий и суспензий.
Энергетический кризис, который охватил в последние годы многие страны мира, привел к необходимости поиска новых нетрадиционных источников энергии. Так, в США с этой целью изучается возможность разведения гигантской ламинариевой водоросли Макроцистис с последующей переработкой ее в метан. Подсчитано, что с площади 400 км 2 , занятой этой водорослью, можно получить 620 млн. м 3 метана.
Гетероконтные водоросли из классов Золотистые, Желтозеленые, Синуровые, Рафидофитовые и Евстигмовые водоросли представленные, в основном, микроскопическими организмами, широко распространены в пресных водоемах всех климатических зон земного шара, но чаще встречаются в умеренных широтах. Есть среди золотистых водорослей виды, которые обитают в морях и соленых озерах, а очень немногие – в загрязненных водах. Максимума развития золотистые водоросли достигают в холодное время года: в планктоне они доминируют ранней весной, поздней осенью и зимой. В это время они играют значительную роль как продуценты первичной продукции и служат пищей организмам зоопланктона.
Некоторые золотистые водоросли, например Уроглена и Динобрион, развиваясь в массовом количестве, способны вызывать цветение воды. Они выделяют альдегиды и кетоны, которые могут придавать воде неприятный запах и вкус, a Уроглена –токсичные для рыб жирные кислоты.
Рафидовые водоросли широко представлены в планктоне пресных водоемов преимущественно с кислым рН, особенно в сфагновых болотах, реже в крупных озерах. В пресных водоемах локальные «цветения» может формировать Гониостомум. Рафидовые водоросли встречаются также в опресненных морских бухтах и лужах на берегу моря, а также в открытом море. При массовом развитии в прибрежных морских водах они вызывают токсичное "цветение" воды. Так, у побережья Канады во время "цветения" концентрация клеток рафидовой водоросли Гетеросигмы может достигать 30 млн. на 1 л. Вспышки численности рафидовых водорослей часто приводят к развитию «красных приливов», с которыми связывают гибель рыб. Причиной таких «красных приливов» могут быть виды родов Хаттонелла, Олистодискус, Гетеросигма и Фиброкапса .
Синуровые водоросли при массовом развитии в пресных водоемах могут придавать воде неприятный запах (Синура ). Феотамниевые водоросли встречаются в стоячих и медленно текущих пресных водоемах, где поселяются эпифитно на нитчатых водорослях.
Евстигмовые водоросли встречаются только в пресных водоемах или в почве.
Желтозеленые водоросли распространены на всех континентах, они обитают преимущественно в пресных водах и почве, а также в наземных, солоноватоводных и морских местообитаниях. Желтозеленые водоросли населяют чистые и загрязненные воды, с различным значением рН: могут обитать как в кислых, так и в щелочных водах. Они встречаются, главным образом, в чистых пресноводных водоемах, реже в морях и солоноватых водах, предпочитая умеренную температуру, чаще развиваются весной и осенью, хотя имеются виды, встречающиеся на протяжении всех периодов года, в том числе и зимой. Чаще всего их можно найти в скоплениях нитчаток и среди зарослей высших водных растений в прибрежной зоне рек, прудов, озер и водохранилищ.
Подавляющее большинство желтозеленых – свободноживущие формы, но встречаются и внутриклеточные симбионты – зооксантелы в клетках простейших. Интересный внутриклеточный симбиоз образуют хлоропласты морского вида Вошерии с моллюском Элизия . На протяжении девяти месяцев этот моллюск способен в культуре к фотоавтотрофной фиксации углекислого газа. Это самый длительный во времени симбиоз такого типа, когда симбиотическая пластида находится в непосредственном контакте с цитоплазмой животного. В природе личинки моллюска питаются нитями Вошерии . В результате фагоцитоза хлоропласты водоросли попадают в цитоплазму эпителиальных клеток моллюска. В ходе этого процесса оболочка хлоропласта становится трехслойной, теряется одна наружная мембрана ХЭС (хлоропластной эндоплазматической сети). Подобное явление служит хорошим доказательством того, что в ходе эволюции в результате вторичного симбиогенеза за счет потери мембран могли возникать хлоропласты с тремя мембранами.
Желтозеленые, золотистые и другие гетероконтные водоросли являются продуцентами кислорода и органических веществ; они входят в состав пищевых цепей. Гетероконтофиты участвуют в самоочищении загрязненных вод и почв, образовании сапропеля, в процессе накопления органических веществ в почве, влияя на ее плодородие. Они используются как показательные организмы при определении состояния загрязнения вод; желтозеленые водоросли входят в состав комплекса микроорганизмов, применяемого для очистки сточных вод.
Контрольные вопросы
- Назовите характерные черты строения бурых водорослей.
- Особенности строения талломов бурых водорослей.
- Как размножаются бурые водоросли? Что такое моноспоры, тетраспоры и зооспоры, изогамия, гетерогамия и оогамия?
- Каковы жизненные циклы у бурых водорослей? Размножение фукусовых и ламинариевых водорослей.
- Назовите характерные особенности и типичных представителей порядков бурых водорослей.
- В каких местообитаниях встречаются бурые водоросли? Каково значение их в природе.
- Хозяйственное значение бурых водорослей.
- Назовите характерные черты строения и типичных представителей золотистых водорослей.
- Какие пигменты и типы питания известны у золотистых водорослей?
- Размножение и экология золотистых водорослей.
- Назовите характерные особенности и типичных представителей желтозеленых водорослей.
- Какие пигменты и типы питания известны у желтозеленых водорослей?
- Как размножаются желтозеленые? Виды полового размножения: изогамия, гетерогамия и оогамия?
- Назовите характерные черты строения и типичных представителей синуровых водорослей.
- Размножение и экология синуровых водорослей.
- Назовите характерные черты строения и типичных представителей феотамниевых водорослей.
- Размножение и экология феотамниевых водорослей.
- Назовите общие черты строения и типичных представителей рафидофитовых водорослей.
- Размножение и экология рафидофитовых водорослей.
- Назовите общие черты строения и типичных представителей евстигмовых водорослей.
- Размножение и экология евстигмовых водорослей.
- Значение гетероконтных водорослей в природных экосистемах.
5.2.4. Отдел Гаптофиты – Haptophyta (Примнезиофиты – Prymnesiophyta)
Одноклеточные, монадные, подвижные, или, редко, колониальные, нитчатые, прикрепленные организмы. Гаптофиты по ряду признаков близки гетероконтным водорослям. Имеется целлюлозная клеточная стенка, иногда с органическими или обызвествленными чешуйками. Все представители примнезиофит имеют характерную органеллу – гаптонему , или "добавочный жгутик".
Клетки с одним ядром, содержащим ядрышко и конденсированные участки хромосом. Хлоропласты по одному-два в клетке, золотисто-бурого цвета, окружены 4 мембранами, из которых 2 собственные. Тилакоиды в группах по три. Пигменты - хлорофиллы а и с , каротиноиды. Митохондрии с трубчатыми кристами. Основным запасным веществом является β -глюкан. Имеют особую органеллу – гаптонему, расположенную между жгутиками (рис. 48). Гаптонема представляет собой нить толщиной, близкой к толщине жгутиков, и различающуюся по длине у разных видов: от 1 до 100 мкм. Основу ее составляют 6-8 серповидно расположенных микротрубочек, окруженных каналом ЭПС.

Рис. 48. Диаграмма строения жгутикового аппарата примнезиевой водоросли Плеврохризис (по: С. Hoek van den et al., 1995): 1 – первый микротрубочковый корешок; 2 – второй микротрубочковый корешок; 3 – третий микротрубочковый корешок; 4 – гаптонема: 5 – дополнительные микротрубочки, отходящие от первого корешка; 6 – дополнительные микротрубочки, отходящие от второго корешка; 7 – базальное тело; 8 – соединение между базальными телами; 9 – соединение между базальным телом и гаптонемой; 10 – соединение между базальным телом и корешком
По-видимому, гаптонема у некоторых видов примнезиофитовых водорослей выполняет роль прикрепительного аппарата.
Чешуйки. Под плазмалеммой у примнезиофит расположен слой цистерн эндоплазматической сети, а сверху от плазмалеммы клетки покрыты одним или несколькими рядами разных чешуек (рис. 49, А ). Органические чешуйки имеют характерную структуру: радиально расположенные фибриллы на внутренней поверхности и концентрически расположенные фибриллы на наружной. Наиболее примитивными чешуйками считают тонкие дискообразные или эллиптические целлюлозосодержащие пластинки. Они встречаются у тех представителей, у которых отсутствуют кокколиты.

Рис. 49. Чешуйки примнезиофит (по: Беляковой Г.А. и др., 2006): А – органическая чешуйка; Б-Д – неорганические чешуйки (кокколиты)
Кокколиты (рис. 49, Б-Д) – кальцинированные неорганические чешуйки; с их помощью регулируется плавучесть клеток и которые выполняют защитные функции. На основе структуры и по месту образования выделяют 2 типа кокколитов: гетерококколиты, которые формируются внутриклеточно (в аппарате Гольджи), и голококколиты, формирующиеся внеклеточно. Голококколиты состоят из правильных ромбовидных и шестиугольных кристаллов, гетерококколиты обладают ромбовидными кристаллами.
Общая характеристика
Клетки обычно свободноплавающие, с двумя жгутиками. Если жгутики заметно неравные, более короткий иногда редуцирован. Иногда более длинный жгутик с тонкими нетрубчатыми волосками. Равные жгутики обычно гладкие, никогда не имеют трубчатых волосков – мастигонем. Если жгутики равные, их, вероятно, больше двух. Размеры их тела обычно не превышают в длину 30 мкм, неподвижные колонии могут достигать 8 мм в диаметре (например, макроскопические колонии Феоцистис). Форма клеток варьирует от округлой до овальной и уплощенной. В жизненном цикле у некоторых представителей могут присутствовать нитчатые, амебоидные, коккоидные и пальмеллоидные стадии.
Пигменты – хлорофиллы а и с , β -каротин.
Большинство видов примнезиофит – фототрофы. Примнезиофитовые способны поглощать бактерии и мелкие водоросли. Фаготрофия осуществляется следующим образом: частичка прилипает к гаптонеме за счет группы сахаров на ее поверхности и передвигается к ее основанию, где расположен центр агрегации частиц. Сформированная крупная частица передвигается вверх к концу гаптонемы, затем гаптонема загибается к заднему концу клетки, где формируется пищеварительная вакуоль, в которой частичка пищи переваривается. Вероятно, содержание фосфора внутри клеток влияет на фаготрофию, и фосфолипиды бактерий используются как источник фосфатов для клеток примнезиофит.
Запасные продукты ассимиляции – парамилон, хризоламинарин.
Бесполое размножение осуществляется путем митотического деления клетки. Половой процесс – анизогамия.